真菌发光基因在植物学研究中的新视角与应用前景
真菌发光基因在植物学研究中的新视角与应用前景
摘要:为探讨真菌发光基因在植物学研究中的应用及其在植物生长、病理学、生态学和分子生物学方面的潜力,介绍了真菌发光基因的基本特征,包括其发现、结构和功能;详细阐述了这些基因在植物学各分支领域的具体应用,突出其在实验技术上的创新,如基因克隆、转录调控和转基因技术等。此外,还分析了这些基因对于理解植物生长机制、病理学特性和生态学关系的意义,并预测了该基因在植物学未来研究中的发展趋势。
生物发光(Bioluminescence,BL)是广泛存在于多种生物体中的自然现象,包括细菌、真菌、鱼类、昆虫和其他海洋生物[1-5]。这一现象由特定基因编码的酶催化的化学反应所产生[6-9]。亚里士多德首次观察到腐烂木材的发光现象,从而区分了生物自主发光与火焰发光,标志着生物发光现象的首次记录[10]。真菌发光基因的系统研究始于19世纪[11],并在20世纪初期通过对特定“发光真菌”种的研究得以深化。科学家们发现这些真菌能发出微弱的蓝绿色光,该现象由特殊酶萤光素酶催化[12]。
随着对真菌生物发光的深入研究,研究人员发现所有已知的生物发光真菌共享相同的发光系统[13],为利用真菌发光基因在植物学生物成像研究[14]及设计可持续发光植物方面的应用开辟了新途径。这些基因作为生物发光报告器,可用于实时监测植物生长和发育过程中的基因表达[15],并用于研究植物对真菌病原体的抗性及植物与真菌的相互作用[16-17]。
虽然在真菌发光基因的研究领域取得了显著进展,但仍面临诸多挑战。首先,需要更深入地探究真菌发光基因的生物化学机制。其次,亟须开发更高效的技术以提取和分离这些基因,并开发高效方法将其引入植物体内。此外,创新用于检测和分析这些基因表达和功能的技术,也是研究的重要方向。总体而言,真菌发光基因在植物学研究中具有广阔的发展潜力,未来的研究可聚焦于植物生长发育、植物病理学、植物生态学和植物分子生物学等多个领域,旨在实现理论和应用层面的更多突破[18-19]。
1真菌发光基因
1.1真菌发光基因的多样性和功能差异
迄今为止,研究者已经发现并鉴定了众多真菌发光基因,这些基因主要源自基生菌门下的不同真菌。基于这些基因编码的萤光素酶的氨基酸序列,可将这些基因分为长链和短链两大类。这两类真菌发光基因在结构和功能上存在显著差异。长链真菌发光基因编码的萤光素酶属于脂肪酰-CoA氧化还原酶超家族,其发光反应需要氧气及胞外长链脂肪酰-CoA作为底物。相反,短链真菌发光基因编码的萤光素酶则属于短链脱氢酶/还原酶超家族,能在无氧条件下催化发光反应。真菌发光基因作为萤光素酶的关键编码基因,构成了真菌发光的分子基础。这些基因的发现和鉴定为深入理解真菌发光机制提供了关键线索。
1.2真菌发光基因特点
真菌发光现象在真菌界中广泛分布。19世纪初,维也纳大学教授Heller首次发现了发光真菌,他通过研究腐烂木材发出的光,证实了其源自真菌发光[11]。2009年,科学家们发现发光真菌中存在酶介导的生物发光反应[12],此后对真菌发光原理研究不断深入[13,20]。2017年,Kaskova等[21]在Neonothopanusnambi真菌中鉴定出萤光素酶可水解咖啡酸,明确了该真菌生物发光生物合成周期,并鉴定了其中涉及的4种酶[13,22],从而确定真菌发光原理借助于咖啡酸循环。咖啡酸循环是所有植物中存在的一种代谢途径,该系统借助其将咖啡酸转化为荧光素(图1),从而使真菌产生发光现象,且真菌发光强度取决于咖啡酸浓度[23]。Mitiouchkina等[24-25]通过农杆菌介导的DNA盒随机位点整合技术,设计了真菌生物发光基因系统。这一系统在真菌发光基因研究中创造了重要里程碑,为真菌发光基因在植物中的应用带来了历史性突破。由于其相较于海洋和陆生生物发光基因的多种优势,发光真菌的发光基因在植物学领域得到了更广泛的应用。
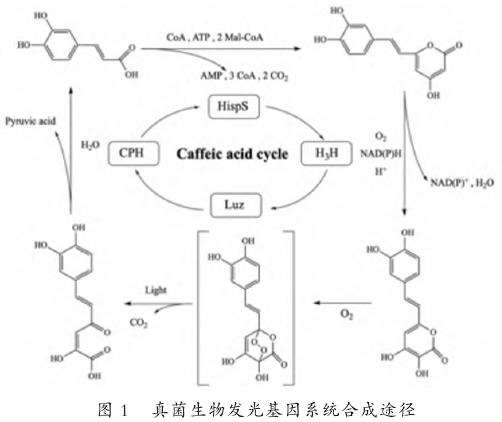
1.2.1真菌发光基因具有裸眼可视性。PurtovKV等[20]的研究表明,真菌的生物发光主要依赖于荧光素。他们鉴定出Neonothopanusnambi真菌的萤光素酶基因属于一个新的蛋白质家族nnLuz[22],并成功确定了完整的荧光素生物合成途径及其涉及的4种酶(nnluz、nnhisps,nnh3h和nncph)。真菌荧光素可以在一系列生物体内表达,使之发出生物亮光[26]。生物发光(BL)的一个重要特点是,其观测无须借助外部光源[25],仅通过咖啡酸循环转化产生的荧光素分子即可实现,因此真菌发光能够在黑暗环境中自行发生,并可被肉眼直接观察到。
1.2.2真菌发光基因具有稳定性。由于真菌发光依赖于咖啡酸循环[24],其产生的光信号极为稳定,并具有高分辨率。因此,在利用真菌发光基因对植物样品进行研究时,可以有效避免加热样品、光源漂移、光散射或背景荧光等干扰因素的影响[25]。这使得研究者能够直接观测植物样品自身的发光强度和持续时间,进而有效地简化对定量信号的评估过程。
1.2.3真菌发光基因具有灵敏度。真菌的发光现象是由酶催化过程产生的,因而对催化反应的稳定状态的扰动反应极为敏感。Strack等[26]在对植物进行创伤处理时发现,真菌生物发光基因能够实现快速地动态测量。此外,Ronzhin等[27]在对发光真菌进行处理后,识别出一种发光刺激剂,即一种低分子量的热稳定化合物,可使真菌发光的光发射水平提升1.5个数量级。利用这一特性,科学家们完成了对真菌生物发光基因的动态测量,并显著提高了其发光系统的灵敏度。
1.2.4真菌发光基因具有高度保守性。通过比较不同真菌谱系(如Mycenaluxenvente、Armillariamelea、Neonothopanusgardneri和Gerronemaviridiulcens)的生物发光基质和转化酶,研究人员发现所有已知的发光真菌共享同一套生物发光系统[13],表明真菌发光转化系统具有高度的保守性。同时,研究发现所有真菌的生物发光(BL)颜色相同。这些真菌发光产生的光均集中在可见光谱的绿色光部分,说明真菌发光的光反应颜色也具有高度保守性。鉴于绿色光能够更好地适应色素植物组织的光学透明窗口[25,28],真菌发光基因的存在为植物生物成像和设计可持续发光植物提供了新的可能性。
2真菌发光基因在植物学中的应用
真菌发光基因在植物学研究中的应用已经取得了显著的进展,以下是一些主要的应用领域。
2.1植物生长和发育研究
真菌发光基因可作为生物发光报告器,用于实时监测植物生长和发育过程中的基因表达[29]。2018年,Kotlobay等[22]展示了真菌中可遗传编码的生物发光系统的潜力,指出真菌生物发光途径可以提供监测基因表达的报告基因。他们发现,毕赤酵母(Pichiapastoris)中含有部分发光基因。该酵母异源表达了N.nam-bi-luz、h3h和hisps3种发光基因,并在含有咖啡酸的培养基中成功实现了这些基因的表达[22]。随后,研究人员在烟草植物中成功引入了真菌发光系统,该系统利用植物中存在的咖啡酸转化为发光物质,从而在无需外部底物的条件下实现了肉眼可见的自发光[23-24]。这一发现为植物成像工具的开发奠定了重要基础。
目前,研究人员已经成功地将Neonothopanusnambi真菌的发光基因(NeoLuc)引入植物中,用于定量分析基因表达[15]。这一方法有助于深入理解植物生长发育的分子机制。Mitiouchkina等[24]的研究也证实了这一点,他们重点研究了真菌发光转基因烟草系的表现,发现该路径的表达无毒性,且不影响植物的生长。此外,Khakhar等[23]还通过瞬时表达法产生了其他自发光植物,包括模式植物拟南芥,观赏植物大丽花、玫瑰、长春花和牵牛花等。
综上所述,在无需添加任何外部底物的条件下,真菌生物发光基因能够在多种植物体内轻松实现表达。更为重要的是,通过真菌发光基因产生的光是肉眼可见的,并且光强优异。这些发现强调了利用真菌发光基因作为生物发光报告基因,实时监测植物生长发育过程中基因表达的可行性与有效性。
2.2植物病理学研究
真菌发光基因在植物病理学研究中主要应用于研究抗真菌病性[30]。通过将真菌发光基因引入植物体内,研究人员可以观察植物对真菌病原体的抗性反应。例如,研究发现,Neonothopanusnambi培养基对引起月季榴莲根腐病的棕榈疫霉(Phytophthorapalmivora)显示出抗真菌活性[31],有助于改善月季榴莲植物的根部腐烂病情,暗示N.nambi培养基可能作为一种农业生物防治剂。最近的研究更加强调了利用真菌发光基因研究植物抗病性的潜力。Mekapogu等[32]特别强调了在观赏作物中研究植物与真菌感染相关的抗性代表性机制。同时,Qiu等[33]指出,玉米的许多研究都集中在真菌病害上,反映出对了解植物抗真菌病原体机制的浓厚兴趣。这些研究成果凸显了人们越来越关注利用真菌发光基因来研究和提升植物的抗真菌病性。
2.3植物生态学研究
真菌发光基因在植物生态学研究中的应用主要集中于探索植物与真菌之间的复杂相互作用。众多研究关注了植物与真菌关联的生态学意义[22,25,32-36]。例如,Xing等[35]强调了植物与菌根真菌之间的相互作用,研究了生命形式对热带森林中兰花菌根网络结构的影响。Galindo-Solís等[36]讨论了植物与内生真菌之间的复杂化学相互作用,指出它们之间关系的多样性。Taudiere等[34]探讨了外生菌根(ECM)共生对于建立植物物种间真菌介导的生态互动关系的潜力。此外,研究人员已利用真菌发光基因研究拟南芥与丛枝菌根真菌(AMF)之间的相互作用[18],通过关联AMF丰度与宿主植物转录,揭示了植物与AMF之间的适应策略,为植物抗逆性研究提供新视角[18]。
同时,通过在植物体内表达真菌发光基因,科学家们也能研究植物生物钟调控机制[37]。研究发现,转入真菌发光基因的植物所发出的光与植物的发育阶段和昼夜节律相关,其发光模式与植物发育阶段的进程、生物钟的调控机制、养分运输和损伤程度紧密相关[25]。因此,通过观测植物的发光模式在时间尺度上的变化,科学家们能够监测和理解植物从萌发到开花的各个阶段的生物调控过程,从而突显了真菌发光基因在生态学研究中的重要意义。
2.4植物分子生物学研究
遗传学和生化学角度对真菌发光系统的特征进行了广泛研究,这些研究主要涉及基因表达模式、基因功能及其与环境的相互作用。例如,Kotlobay等[22]从真菌中成功鉴定出萤光素酶及其在真核生物中荧光素生物合成中的作用,为理解真菌发光系统的遗传和生化特性提供了重要见解。同样,Zhang等[38]在植物原生质体中通过瞬时基因表达技术,深入探究了基因功能、蛋白质的亚细胞定位以及蛋白质间的相互作用。此外,Richards等[39]通过对拟南芥天然品种的全基因组表达模式评估,研究了其在整个生命周期中的转录变化及其与环境波动的相关性。Schenk等[40]的研究则聚焦于拟南芥接种真菌病原体后选定基因表达模式的变化,强调了理解基因表达对环境刺激反应的重要性。这些研究共同揭示了真菌发光基因在揭示植物分子生物学中基因表达模式、基因功能及其与环境相互作用方面的关键作用。进一步,Tong等[41]描述了参与真菌生物发光的hispidin3-hydroxylase的特性,为理解发光途径中这一关键酶的底物结合和反应性提供了深入洞见。这些研究不仅增强了我们对真菌发光基因生化机制和特性的理解,而且强调了其在植物分子生物学领域中,特别是在揭示基因表达模式、基因功能与环境相互作用方面的核心作用。
2.5植物环境污染测定研究
重金属和有机污染物对生态系统和人类健康产生重大影响,成为全球广泛关注的问题。尽管传统的化学吸收方法,如原子吸收光谱法和原子发射光谱法,用于测定环境中的重金属和有机污染物浓度时反应灵敏,但这些方法往往耗时较长。生物发光技术则成功克服了这些缺陷。自2002年起,生物发光真菌Armillariamellea和Mycenacitricolor已被用于开发一种测定环境重金属和有机污染物浓度的新方法[42]。该方法的原理是:真菌的发光强度会随着环境中特定重金属(如铜和锌)和有机化合物(例如氯酚)的浓度而变化。通过测定这些真菌的生物发光特性,研究人员能够推断环境污染物的浓度。
近年来,研究还发现,酚类化合物会抑制发光真菌Gerronemaviridilucens的发光能力,这进一步证实了利用发光真菌评估环境污染物毒性浓度的可行性[43]。综上所述,真菌发光基因在植物学研究领域的应用已取得显著进展,它不仅为我们提供了一种强有力的研究工具,还有助于深入理解植物的生长、发育、病害抵抗能力以及与真菌的相互作用等重要生物学问题。
3研究方法与技术
在植物学研究中,应用真菌发光基因涉及一系列的研究方法与技术,包括从真菌中提取和分离发光基因、基因克隆与表达、转基因技术的应用,以及相关的检测与分析方法。
3.1真菌发光基因的提取和分离
真菌发光基因的提取与分离是实验的初步阶段。通常,此过程采用生物化学技术,例如超高效液相色谱联合飞行时间质谱(UPLC-Q-TOF-MS)技术,以从真菌样本中准确分离出目标基因[44]。
3.2真菌发光基因的克隆和表达
真菌发光基因的克隆与表达是实验的核心部分。此过程通常采用反转录聚合酶链反应(RT-PCR)和快速扩增cDNA末端(RACE)技术,以获取基因的完整开放阅读框架(ORF)[45]。继而,通过构建原核表达载体并在大肠杆菌中诱导基因表达,实现基因的功能研究[45]。
3.3真菌发光基因在植物中的转基因技术
将真菌发光基因引入植物体,是通过转基因技术实现的。与传统育种技术相比,基于遗传转化的分子育种方法可以有效缩短育种周期,简化繁复程序,并实现目标性状的定向改良[46]。此外,通过运用CRISPR/Cas9基因编辑技术,可以将外源基因精确地插入植物基因组[47-48]。由于真菌荧光素的合成底物咖啡酸在所有植物中均有存在,因此转基因技术在提高植物的抗虫害、抗逆性、抗病性等方面发挥着关键作用,同时也用于调节植物的本质特性、开花周期及修复植物[49]。
目前,植物转基因技术的研究重点在于提高标记基因的转化效率、转基因植物组织再生,以及探究基因功能调控机制。通过基因工程技术,研究人员能将真菌生物发光途径酶编码基因导入烟草核基因组[24],设计出能促使植物体内由咖啡酸到荧光素分子转化的质粒序列,从而实现无需外加化学物质的植物自发光[23]。
3.3.1基因枪法。基因枪法是通过高速将附着外源遗传物质的金属颗粒注入细胞受体的方法,实现目标DNA传递至目标生物体内[50]。此技术主要应用于农作物(如小麦、豆类)及某些果树(如柑橘)的遗传转化[51]。基因枪法具有快速、简便、高效的特点,并能够实现对花粉、胚胎、分生组织和形态细胞等全能组织的直接转化。然而,该技术需要特殊设备,成本较高,且仍处于发展初期[52],因此在植物学研究中尚未广泛应用。
3.3.2农杆菌介导法。农杆菌介导法是植物转基因技术中的首选方法,自大约40年前首次成功应用以来,一直被广泛采用[53]。根癌农杆菌可将DNA分子传递到植物细胞中,并将外源基因整合入宿主基因组[54]。这一方法简便、经济且高效,具有基因沉默现象少、转化周期短和能转化较大片段基因等优势,因而受到科研工作者的青睐[55]。它对于研究基因功能及培育转基因植物具有重要意义。然而,根癌农杆菌只能感染有限的农作物品种和基因型,其残留物可能引起植物形成冠瘿,进而影响产量[56]。在应用过程中,农杆菌株系和菌液浓度对转化效率有显著影响[21]。研究显示,EHA系列菌株相比GV3101、LBA4404等具有更佳的转化效果[57]。赵亭亭[58]的研究发现,EHA105菌株更易于激活且适用性更广。至于菌液浓度,低浓度(0.4~0.6mg/L)的菌液可减轻植物性毒素的影响。农杆菌介导法常与基因枪法结合使用,以提高转化效率[59]。BouanamaB和BidneyD等[60-61]的研究表明,结合微粒轰击和农杆菌介导转化的方法比传统基因枪法的转化率高出至少100倍。
3.4真菌发光基因的检测和分析
对真菌发光基因在植物中的表达与功能进行检测与分析,通常需采用多种分子生物学和生物化学技术。例如,碱基猝灭探针技术可用于检测基因特定突变[62],变性梯度凝胶电泳(DGGE)技术则用于分析基因表达模式与菌群多样性[63]。这些技术的应用使研究人员能更深入地理解真菌发光基因在植物生长发育中的作用,为植物病理学和植物生理学的研究提供了重要工具。
4真菌发光基因在植物中的未来发展方向
真菌发光基因在植物学研究的应用已取得显著进展,但该领域仍面临多项挑战与研究机遇。
4.1分析检测
真菌发光基因的主要优势在于其提供了一种非侵入性的研究手段,使科学家能够清晰地观察和跟踪植物的生长发育过程,尤其是在根部等难以观察的区域。利用发光基因,研究人员可以实时监测植物的生理状态,包括生长、病理反应和环境应激反应,这对于理解植物如何响应不同环境条件(例如干旱、盐胁迫或病原体攻击)至关重要。例如,通过利用真菌发光基因标记植物的抗病基因,可以筛选出具有强抗病性的植物品种。然而,某些植物具有自发荧光的特性,实际研究中需通过显微镜验证真菌发光基因在植物体内的表达,以避免误判[64]。真菌发光基因在监测环境污染物苯酚方面也具有巨大的应用潜力。例如,真菌N.nambi的胞外氧化酶是含FAD的蛋白,能催化苯酚化合物以检测苯酚的存在,此类氧化酶亦出现在新型发光真菌Oth-opanusnambi中,有助于水介质中苯酚的检测[65-66]。发光真菌的分析应用潜力巨大,未来需要更多研究以深入探索和发现。
4.2成像与传感
使用β-葡萄糖苷酶处理发光真菌后,Ronzhin等[27]从N.nambi的菌丝体中得到了发光刺激剂,该化合物是低分子量、热稳定,能使光发射水平提升1.5个数量级,为开发真菌发光探针和传感器提供了可能。科学家对咖啡酸循环转化为荧光素的机制进行研究,发现咖啡酸通过2种酶转化为发光前体,再由第3种酶氧化生成光子,然后该酶将氧化分子转换回咖啡酸,重新开始循环[24]。此外,有研究探讨聚集诱导发光体系中咖啡酸的作用[67]。目前,对咖啡酸转化为发光体的具体机制尚未完全明了,需要更多研究进行探索。
4.3活性氧的产生与应用
真菌发光是自氧化过程[68],分子氧是真菌生物发光的主要组分[22]。Kobzeva等[69]发现,N.nambi在电离辐射刺激下会产生活性氧(ROS)。这一特性可用于研究ROS在各类生物过程中的作用,并开发基于ROS生成的新型治疗策略。然而,真菌发光基因技术也面临挑战,如发光信号的强度和稳定性可能受多种因素影响(例如植物的发育阶段、环境条件和基因表达水平变化)。此外,将真菌发光基因有效整合入植物基因组,而不影响植物的正常生长发育,也是重要研究方向。总之,尽管真菌发光基因在植物学研究应用前景广阔,但仍需更多研究解决现有挑战,充分挖掘其潜力。随着研究深入,可期待在植物生物学、农业科学乃至生态研究等领域取得更多突破。
5总结与展望
真菌发光基因在植物学研究中的应用已经取得显著进展。这些基因的发现与研究不仅揭示了生物发光的神秘面纱,还为我们提供了一种新颖而强大的研究工具,用以探索植物学中的各种问题。在植物生长发育的研究中,真菌发光基因作为生物发光报告器,可用于实时监测植物生长发育过程中的基因表达[24,27]。在植物病理学方面,真菌发光基因在提高植物抗真菌病性方面已取得显著成果[31]。在植物生态学研究中,它们在探究植物与真菌之间的互作关系方面具有重要应用价值。在植物分子生物学领域,真菌发光基因在研究基因表达模式、基因功能以及基因与环境相互作用方面展现出广阔的应用前景。
然而,尽管真菌发光基因研究已取得显著进展,该领域仍面临多项挑战。例如,在分子层面上,咖啡酸转化为荧光素分子的具体机制尚不清晰,未来需要更深入理解真菌发光基因的生物化学机制,并开发更有效的提取和分离方法。在植物表达方面,真菌发光基因转入植物后的稳定性和表达效率尚不明确,需开发更有效的转基因方法。此外,真菌发光基因对植物的安全性也需进一步研究,以确保其表达和功能的安全性。总体来说,真菌发光基因在植物学研究中具有广阔的发展潜力。未来研究可以集中在植物生长发育、植物病理学、植物生态学和植物分子生物学等领域,以期在理论和应用层面取得更多突破。